
After a small twitter discussion with the simulation theologists (AKA, ghost in the cloud programmer followers) got me thinking of Richard Dawkins and a video on the evolution of the eye. I thought: Gee, someone must already have covered this and indeed, I found a post here in steemit. It consisted of a link to that video and nothing else.
So I think I can contribute a little more with the molecular biology of it for the platform.
Evolution is highly conservative: creates new structures based on random changes and repurposes old and highly conserved structures for new uses.
The Eye
Th evolutionary perceived history of the eye is kind of intuitive. You begin with the idea of life starting in the ocean, some self-replicating structures with a lipid envelope start to gain energetic benefits by being closer to the sunlight, this energetic benefit starts a self-sustaining loop of finding that light. Later specialization appears and the portions that react to light concentrate and you get the first photoreceptor linked to some kind of processing power and movement effectors.
Early fossils date the oldest prototypical eyes to the Cambrian period where the eye-spots, the first units of photoreceptors known appeared. This spots did provide sensitivity but lacked more than a couple degrees in perception, as direction localization seems to have been a rare development but is the responsible for 96% of the living species.¹
The Platynereis dumerilii is considered a living fossil. Is a polychete worm of the same family as earthworms. Possesses prototypical pigment-cup eyes.

The following is a representation of the larval eye at 24 hours as seen traced by E.M. 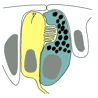 In yellow the Rhabdomeric photoreceptor cells, in green the pigment cells. These two cells are the simplest possible visual system. It senses light, direction, and color. It couples directly to other cells.
In yellow the Rhabdomeric photoreceptor cells, in green the pigment cells. These two cells are the simplest possible visual system. It senses light, direction, and color. It couples directly to other cells. At this stage, it has no lens, no brain, no nerves. Only a single cell retina.
Here at 72 hours, the proto-adult pigment cup eye  the change in the structure has led to directionality enhancement, giving it the ability to locate. Neves will start to grow towards the rhabdomere due to its surface markers.
the change in the structure has led to directionality enhancement, giving it the ability to locate. Neves will start to grow towards the rhabdomere due to its surface markers. At this stage no lens, single cell retina, single optical nerve and no brain
Here, the adult pigment cup eye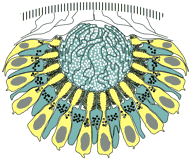 Is the result of a duplication of the larval eye parts.
Is the result of a duplication of the larval eye parts. It has no lens, a multi-cell retina, multiple nerves and a simple brain²
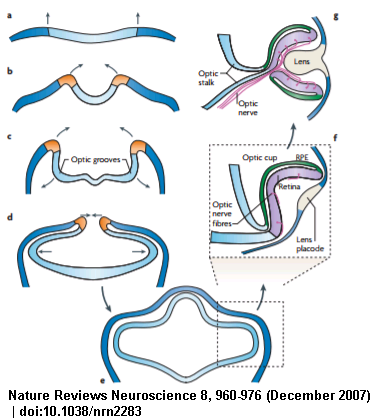
The stereotypical progression in embryonic development in bilaterians gives an idea of the evolutionary progression from the Urbilaterians. This structures at each stage can still be found today.
In fact, a cellular view of the Ascidian, Hagfish eye, Lamprey, and gnathostomes. Reveals high diversity in those closely related organisms but all seem to clearly have evolved from two types of cells. The Rhabdomeric and ciliary photoreceptors. In vertebrates, this correlates to Rods and Cone cells respectively.
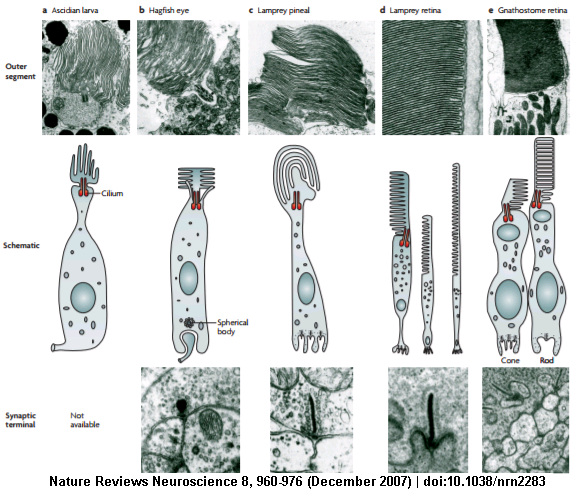
Each of these terminates in a single synaptic terminal, where the retina meets a nerve ending. All sensory tissues have this types of terminals, suggesting that complex eyes could have evolved from a single eyespot. It doesn't require the growth of new neural pathways in development.³
The photoreceptor cells are special, they contain molecules called opsins. Ciliary photoreceptors contain a broad diversity of these, the prototypic being Rhodopsin, derived from vitamin A. This provides a test for evolution.
Assuming the theory of common ancestor is true, we can use comparative genomics. If we examine some of the data compiled in homologene, we see a pattern of genetic similarity that matches the stages of eye complexity development. One should expect Statistical relatedness among all animals tested. Highly conserved genes from the simplest visual systems to the more complex eye. Temporal identification of the genes in evolutive time (ancient to most recent) and a plausible route for them to have alternative functions which could lead to changes to new purposes.
In the case of humans vs Chimpanzee, there's a nearly identical protein sequence for most eye-related genes. Wich makes an initial case for the hypothesis. In a comparison with other mammals, there is a difference but mostly in genes related to low-light perception. In the case of humans, for instance, genes like PIN2 IP and Opsin 4 are more recent and have weaker selection than genes like PAX6 (the homeobox gene that determines where eyes are formed). Genes like rhodopsin, opsin 1 and transducin are highly conserved and makes them more ancient genes.

Transducin, the protein that transmits the signal from the opsins, is highly conserved in Humans and fishes, yet a highly similar structure can be found in plants. Opsin and transducin combined are a minimal visual system. Another part of the simple visual system if GNBP, the signaling molecule involved in extracellular signaling pathways, including rhabdomeric cone cell photosensitivity. The change in statistical variety correlates with changes in morphology and archeological evidence.
If someone examines a gene like PAX6 (part of the Eye disc development genes), it will find that is highly conserved in vertebrates but less conserved in invertebrates, suggesting a predecessor gene for bilaterians that was modified before vertebrates and invertebrates parted evolutionary pathways as there's a big difference in the morphology of the disc structure between vertebrates and invertebrates like some flies.
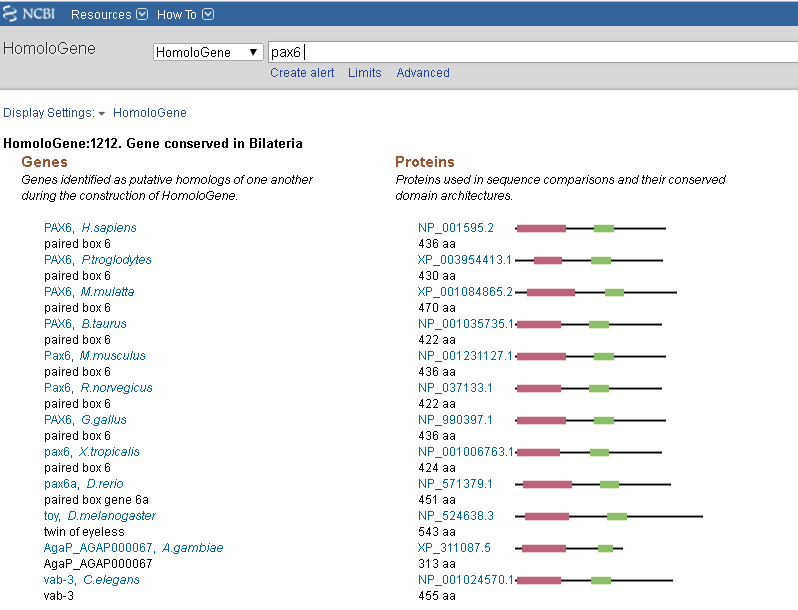
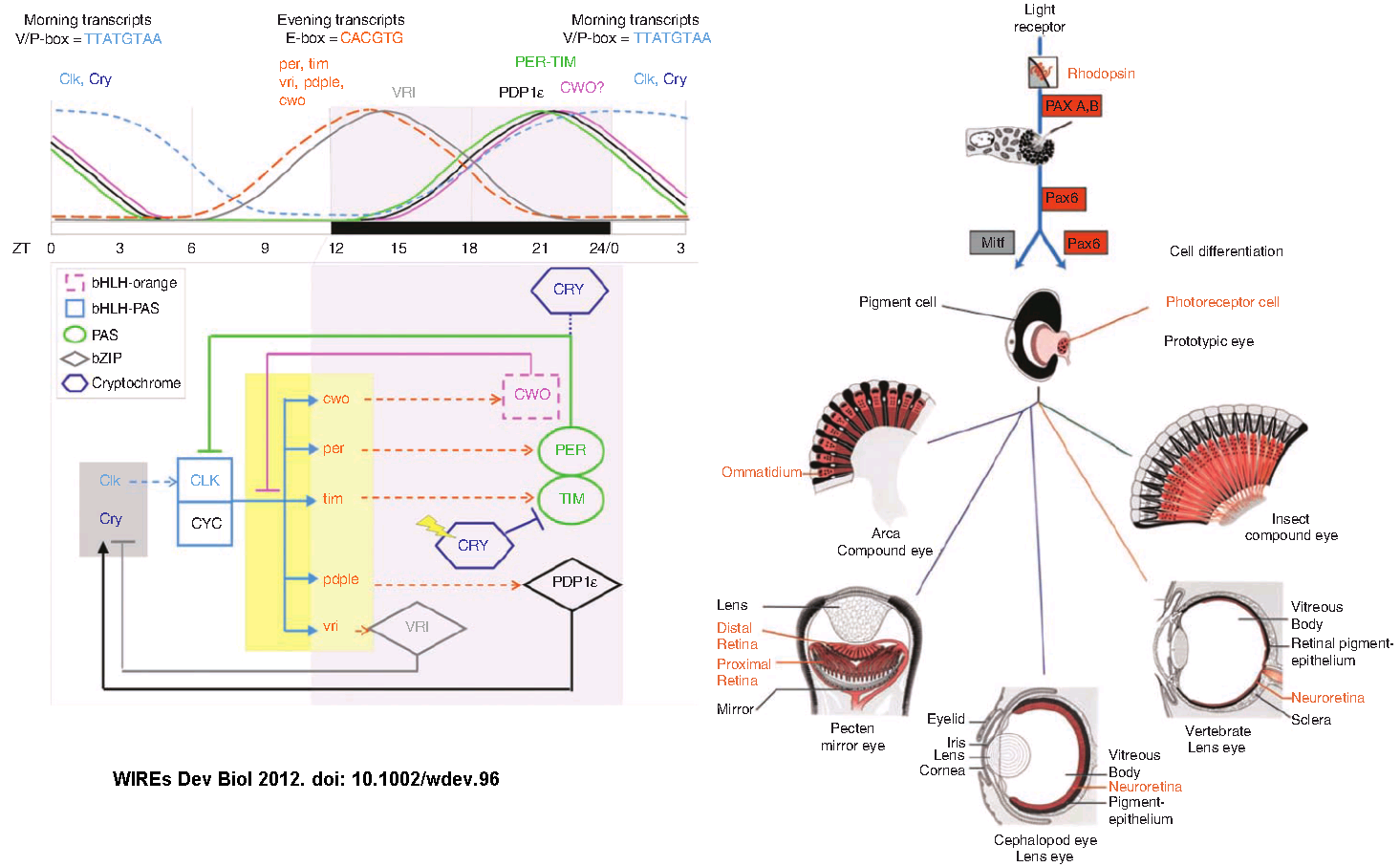
The plausible alternative uses for genes can be seen for instance in opsins. Which is correlated with circadian rhythms in single cell organisms and blue-green phototaxis in algae. Transducin and GNBP are part of cascades in signal transduction. Atonal, PAX6, FOXA2 is part of the basic helix-loop-helix transcription factors part of many sensory systems. Cryptochromes are found in many animals and are related to bacterial photolyase, which repairs ionizing radiation-induced DNA damage using light energy.⁴
How this all mumbo-jumbo relates? One plausible scenario coming from the circadian rhythms observed in Drosophila and cyanobacteria. In the genome of an early heterotropic Eukaryote, a cryptochrome became fused with a transmembrane domain containing a transducin binding site. Producing an opsin which increased the fitness of a photosynthetic cell. The evolution of the specialization in eye structures came from the transposition of an opsin gene with a basic helix loop helix promoter sequence from another gene. These would be selected for and inherited. ⁵
The part of the eye that focuses light onto the retina is composed ot three parts: The capsule, epithelium, and fibers.
The lens capsule is a basement membrane, a type of structure common in tissues that adhere to other tissues. The lens epithelium is simple and cuboidal, it regulates the exchange of ions, nutrients, and fluids, (this same type of epithelium is in the kidney and thyroid).
The lens fiber cells derived from the epithelium and are an anucleated, without organelles long and thin specialization of cells. They are packed tightly and use gap junctions to generate layers (laminae). Is composed mainly of proteins called crystallins. They refract light in the lens. They are not special. They have evolved from other genes (like seems to be the case with crystallin alpha B, found for other purposes in bacteria), according to the protein sequence similarities. These proteins originally were part of the embryonic heart and kidney in vertebrates. ⁶
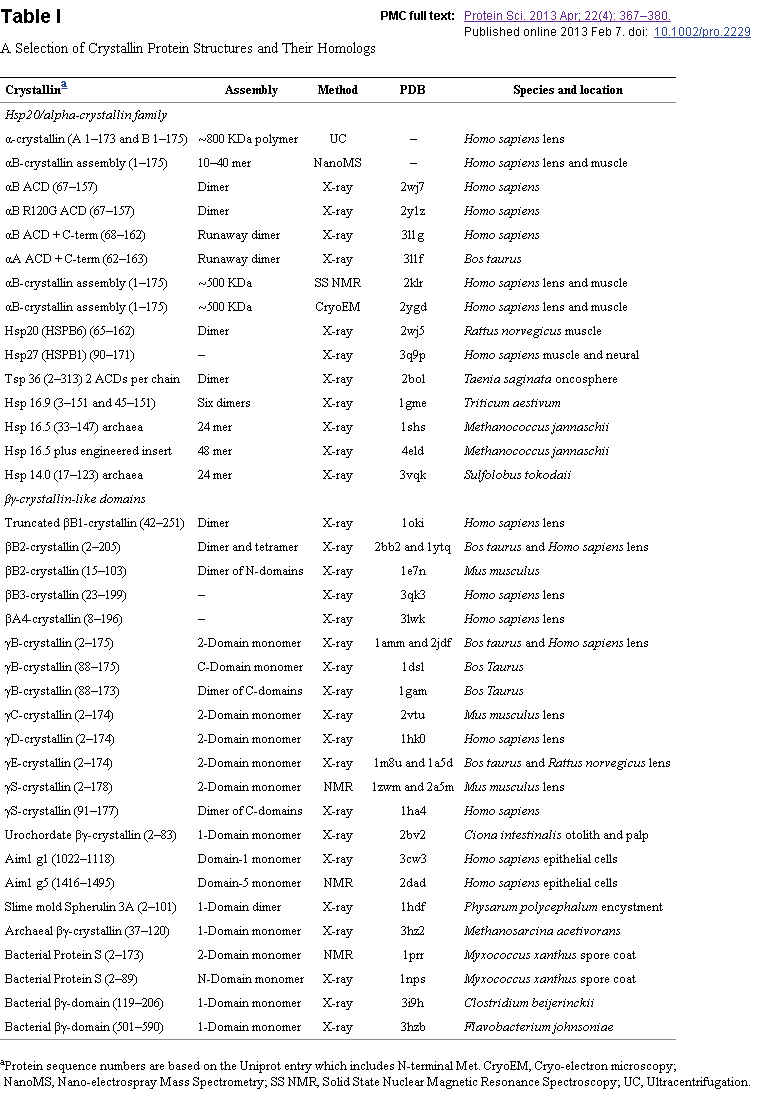
The genetic homology of these structures shows a similar probable origin. The fact they have multiple uses differing from the specialization in a particular tissue indicate that even in a single organism the repurposing of a structure is the norm. A form of biological economy of available materials.
References:
1 M F Land, and R D Fernald. The Evolution of Eyes. Annual Review of Neuroscience Vol. 15:1-29
5 Walter J. The evolution of vision. GehringWIREs Dev Biol 2012.
https://www.ncbi.nlm.nih.gov/homologene
Images sourced, taken from the referenced articles. Introduction image sniped form the Richard Dawkins video for public education linked.
