Verziju ovog članka na engleskom jeziku možete pročitati ovde.
U prethodnom Evolucija 101 članku imali smo priliku da naučimo nešto više o učestalostima alela i genotipova, kao i da se u hipotetičkoj Hardi-Vajnbergovoj populaciji učestalosti alela ne menjaju kroz generacije.
Međutim, u realnim, prirodnim populacijama tako nešto se skoro i ne može očekivati, i uvek dolazi do promene u učestalostima alela. Osnovni evolutivni mehanizmi koji mogu uzrokovati promene u učestalostima alela su sledeći:
- Genetički drift
- Prirodna selekcija
- Mutacije
- Migracije (ili protok gena)
Genetički drift i prirodna selekcija predstavljaju dva najznačajnija procesa koji dovode do evolutivnih promena, i zbog toga ćemo ih detaljnije pojasniti u ovom i narednom članku.
Genetički drift ili bacanje kockica
Po definiciji, genetički drift predstavlja evolutivni mehanizam koji dovodi do nasumičnih (slučajnih) promena u učestalostima alela u populaciji kroz generacije.
Do promene u učestalostima alela dolazi zbog "greške uzorka" tokom procesa uzorkovanja onih alela iz genskog pula koji će učestvovati u formiranju naredne generacije.
Efekat genetičkog drifta je posebno naglašen u malim populacijama.
Genetički drift, greška uzorka, genski pul, slučajni događaji, male populacije - zaista, o čemu ti govoriš? To uopšte ne zvuči kao proces evolucije!

Genetički drift je stohastički proces, isto kao bacanje kockica, autor ejaugsburg, CC0 1.0
Nasumično uzorkovanje u genetici
Da bi lakše razumeli stohastičku prirodu genetičkog drifta, prvo ćemo objasniti koncept slučajnog uzorkovanja na primeru klikera u činiji.
Zamislite da imate činiju sa 100 plavih i 100 crvenih klikera - ukupno 200 klikera, i da imate zadatak da izvučete 100 klikera zatvorenih očiju i da ih prebacite u drugu činiju. Nakon što izvučete 100 klikera, treba da prebrojite koliko ima plavih i crvenih klikera, izračunate njihov odnos, dopunite klikere u novoj činiji do 200 tako da odnos izvučenih plavih i crvenih klikera ostane nepromenjen, i da ponovite izvlačenje.
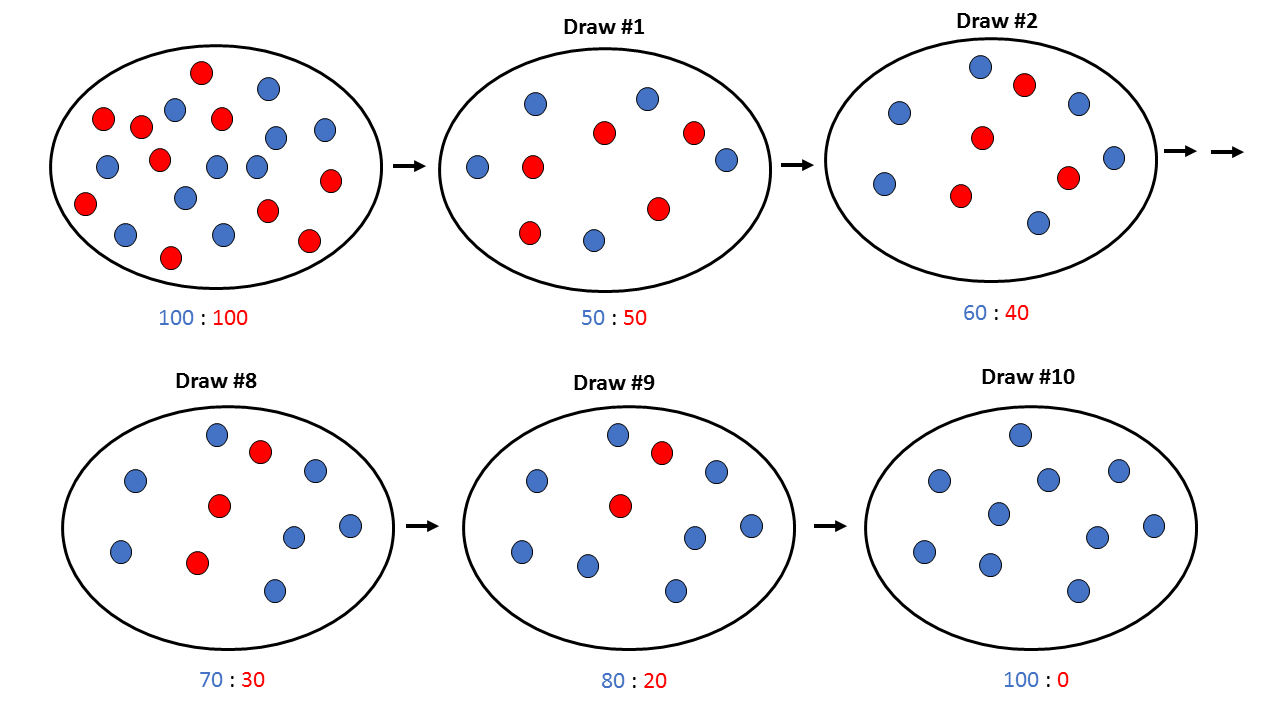
Nakon što ste ponovili izvlačenje/dopunu klikera nekoliko puta (u ovom primeru 10), primetićete da su vam u činiji ostali samo klikeri jedne boje (u ovom primeru plavi)! Crveni klikeri nisu uspeli da se zadrže tokom procesa izvlačenja, što je posledica slučajnosti, ili greške uzorkovanja.
Isto se dešava i sa genskim alelima u populaciji - jednostavno, zbog greške uzorkovanja, neki od alela će biti "izdriftovani" iz populacije (drugim rečima - izbačeni) i nestaće zauvek.
Pogledajmo sada šta se dešava ako umesto 200 počnemo izvlačenje sa ukupno 20 klikera (10 plavih i 10 crvenih), izvlačimo 10 i prebacujemo ih u novu činiju.
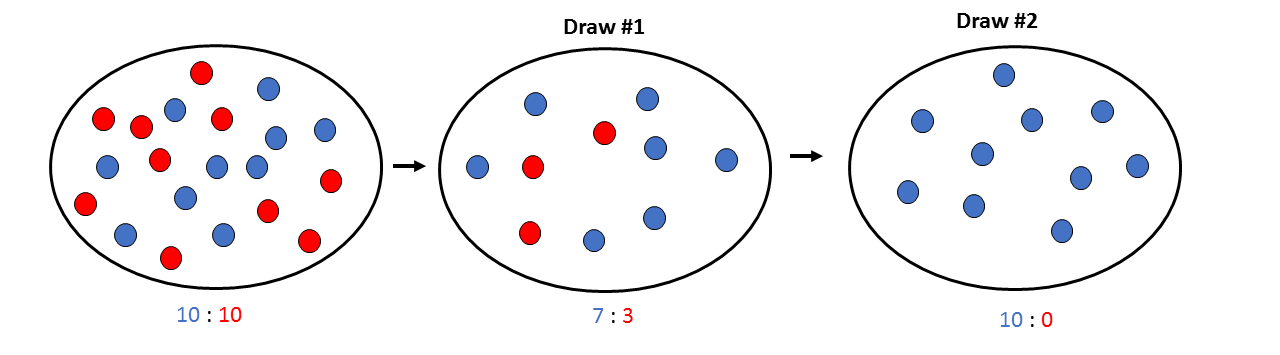
Primećujete kako nam je brzo nestalo crvenih klikera, i to nakon samo dva ciklusa izvlačenja!
Isto se dešava u malim populacijama - genetički drift ispoljava drastičniji efekat u poređenju sa velikim populacijama. Dobar primer za dejstvo genetičkog drifta u malim populacijama su ugrožene vrste, jer gubitak alela uzrokovan genetičkim driftom predstavlja veliki problem za genetičku raznovrsnost ovakvih populacija.
Genetički drift i učestalosti alela
Sada ćemo objasniti genetički drift na konkretnom primeru prirodnih populacija.
Pretpostavimo postojanje male populacije zečeva od 8 braon i 2 bela zeca (10 zečeva ukupno), čiji aleli B i b imaju jednaku početnu učestalost (p = q = 0.5).

Genetički drift u malim populacijama, autor OpenStax College, CC BY 4.0
Dalje možemo pretpostaviti da će se iz prve generacije (F1) razmnožavati samo 5 zečeva, jer će ostali završiti kao plen lovaca (nasumični događaj, jer lovci ne biraju zečeve koje će loviti na osnovu boje krzna).
Tada ćemo imati promenu u učestalostima alela u F2 generaciji (p = 0.7, q = 0.3). Ako se iz nekog razloga u F2 generaciji razmnože samo 2 zeca nosioca BB genotipova, u F3 generaciji alela b više neće biti (q = 0), i jedini preostao alel će biti B (p = 1).
Kada (kao u ovom primeru) učestalost jednog alela postane 1, to znači da je taj alel fiksiran u populaciji, i da sve jedinke imaju isti taj alel za određeni genski lokus. Drugi alel, alel b, je izbačen iz populacije (izgubljen zauvek), drugim rečima njegova učestalost je sada q = 0.
Fiksaciija jednog i potpuni gubitak drugog alela se može smatrati ekstremnim primerom. U suštini, osnovno pravilo kod delovanja genetičkog drifta je da aleli koji imaju male početne učestalosti imaju veću šansu da nestanu u malim populacijama, u poređenju sa alelima većih učestalosti.
Primer sa populacijom zečeva savršeno ilustruje prirodu efekta genetičkog drifta na populacije - genetički drift svojim delovanjem smanjuje genetičku varijabilnost populacija.
Koja je verovatnoća fiksacije alela?
Dejstvo genetičkog drifta je najbolje analizirati na primeru neutralnih alela.
Neutralni aleli su oni aleli koje prirodna selekcija ne "prepoznaje" (više o prirodnoj selekciji u narednom članku), i za koje možemo očekivati da imaju učestalosti predviđene Hardi-Vajnbergovom jednačinom:
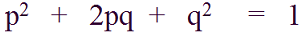
Međutim u praksi, u svakoj generaciji postoji šansa da se učestalost određenog alela poveća, smanji ili da ostane ista, isključivo kao posledica genetičkog drifta, na način na koji je prethodno objašnjeno.
U populaciji veličine N postoji ukupno 2N alela na svakom lokusu. Obzirom da je proces genetičkog drifta potpuno nasumičan, svaki od tih 2N alela će imati podjednaku šansu da bude "odabran", ili fiksiran.
Iz toga sledi da se verovatnoća fiksacije bilo kog alela u populaciji može odrediti na sledeći način:
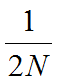
Na isti način se može izračunati i verovatnoća da će bilo koji alel biti izgubljen iz populacije pod dejstvom genetičkog drifta:
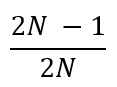
Kako možemo izmeriti genetički drift?
Da bismo to uradili, potrebno je da prvo definišemo efektivnu veličinu populacije (Ne).
Efektivna veličina populacije predstavlja onaj broj jedinki u populaciji koji se razmnožava i koji će proizvesti potomstvo, tj. jedinke naredne generacije.
Drugim rečima, efektivna veličina populacije predstavlja veličinu idealne Hardi-Vajnbergove populacije koja bi "osetila"
isto dejstvo genetičkog drifta kao i posmatrana, realna populacija.
Kao što možete da pretpostavite, Ne vrednosti su obično niže od stvarnog broja jedinki u populaciji koje se ukrštaju međusobno (N):
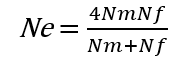
Nm - broj mužjaka
Nf - broj ženki
Na primer, u populaciji koja ima 20 mužjaka i 20 ženki koji su reproduktivno zreli, efektivna veličina populacije bi bila 40, ili jednaka stvarnoj veličini populacije.
Međutim, zamislite populaciju od 100 jedinki, sa 96 ženki i samo 4 mužjaka (haremski oblik poligamije kod životinja) - Ne u takvoj populaciji bi bila 15.36, što znači da takva populacija, iako je njena stvarna veličina 100 jedinki, trpi jednak efekat genetičkog drifta kao i populacija od 15 jedinki!
Sada možemo izračunati koliki efekat genetičkog drifta možemo očekivati nakon jedne generacije (važi za diploidne organizme):
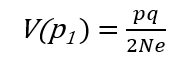
V(p1) - varijansa učestalosti alela
p, q - učestalosti alela na posmatranom genskom lokusu
Ovo suštinski znači da efekat genetičkog drifta zavisi od genetičke varijabilnosti populacije (p, q - učestalosti alela) i od efektivne veličine populacije.
Ako su p = q = 0.5, Ne = 50, i V(p1) = 0.0025, tada će promena učestalosti alela u svakoj generaciji biti:
p = (0.0025)0.5 = 0.05, ili 5%.
U maloj populaciji od samo 5 jedinki efekat drifta će biti veći:
p = q = 0.5, Ne = 5, V(p1) = 0.05
p = (0.05)0.5 = 0.22, ili 22% slučajnih promena učestalosti alela u svakoj generaciji!
Efekat osnivača i uskog grla
Inicijalno veliki broj jedinki u populaciji se može drastično smanjiti prolaskom kroz usko grlo i efektom osnovača.
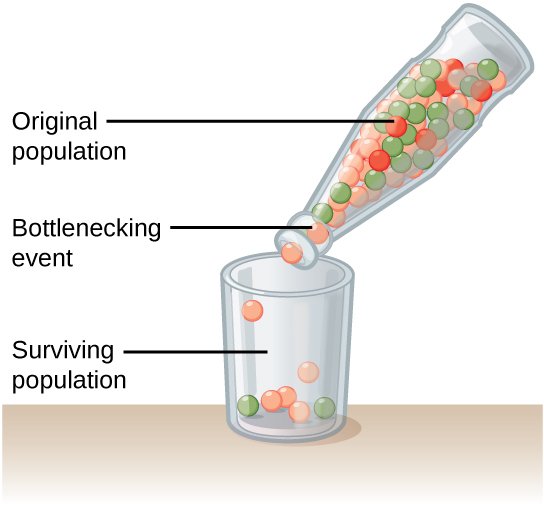
Prolazak populacije kroz usko grlo, autor OpenStax College, CC BY 4.0
Do prolaska populacije kroz usko grlo obično dolazi nakon prirodnih katastrofa, koje ubijaju veliki procenat jedinki u populaciji po principu slučajnosti.
Nakon takve katastrofe, struktura populacije se obično drastično izmeni, jer preživele jedinke postaju nosioci celokupnog genskog pula koji je sada značajno redukovan u poređenju sa inicijalnom populacijom.
Drugi scenario u kome bi populacija "osetila" snažan efekat genetičkog drifta je slučaj gde samo mali deo inicijalne populacije migrira i naseli novu teritoriju, noseći samo deo alela iz originalne populacije. Drugim rečima - te jedinke koje su migrirale obično nisu reprezentativan uzorak cele populacije. Ova pojava se naziva efektom osnivača.
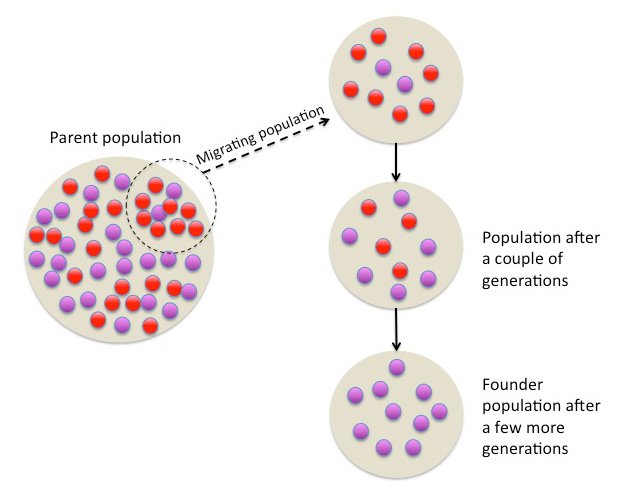
Efekat osnivača, autor Tsaneda, CC BY 3.0
Danas smo naučili šta su slučajni događaji u procesu evolucije i na koji način oni utiču na promene u učestalostima alela. U sledećem članku ćemo videti šta je prirodna selekcija i kako ona menja strukturu populacije.
Literatura
Mechanisms of Evolutionary Change
Evolution, Mark Ridley, 3rd Edition
Effective population size and patterns of molecular evolution and variation
