I will introduce some of the basic concepts that are thought to explain why plants (your salad) can survive in their natural environments.

Figure 1. A resistant (Right) and diseased tomato leaf (left) both inoculated with the bacterial pathogen Pseudomonas syringae (From: Freeman and Beattie (2008)). Image source
Plants are continuously challenged by a diverse microbial community members, some of which can cause disease (Boller and Felix, 2009; Jones and Dangl, 2006; Dodds and Rathjen, 2010). In most cases, infection is limited because of physical (e.g. cell walls) and/or chemical barriers that block the pathogen from gaining access to the host interior (toxins and other secondary metabolites) (Huitema et al., 2003; Schulze-Lefert and Panstruga, 2011).
Not surprisingly, some microbes have acquired and evolved enzymes that help degrade physical barriers or detoxify their host environment. Given the high-end stakes of plant-pathogen interactions (life vs death), it is not surprising that plants have adopted additional defensive strategies to help keep pathogens at bay. Genetic, biochemical and functional studies in model plants such as Arabidopsis thaliana and work in an increasing number of crop species have firmly demonstrated mechanisms of adaptive immunity that fend off microbial attacks (Jones and Dangl 2006). These include local induced responses (site of infection or attack) and systemic responses that are induced by hormones that originate from infection sites (Figure 1). These signals travel throughout the plant, alerting distant tissues of pathogen presence and thereby priming the plant for a faster and stronger response.
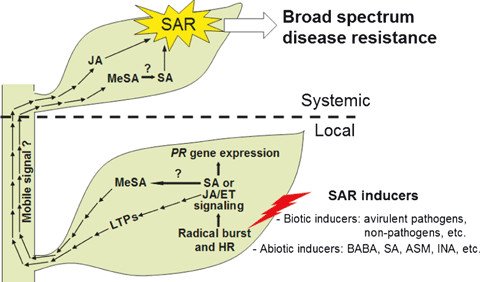
Figure 2. Illustration of local defense responses and their systemic effects on plants (from Choi & Kook Hwang, 2011). Image Source
Early genetic studies have identified a great number of mutants in Arabidopsis, that are either unable to mount systemically induced responses or have lost the ability to limit the expression of defense associated genes (constitutive defense mutants with enhanced resistance). These advances in combination with subsequent biochemical and functional genomics studies have facilitated both the dissection of distinct defense signaling pathways, as well as the identification of gene complements whose expression is defense associated. Perhaps not surprisingly, many of the proteins encoded by defense-associated genes have enzymatic functions (e.g. lytic enzymes that break down pathogen cell walls, produce anti-microbial metabolites) or act as inhibitors of pathogen-encoded enzymes (Tian et al., 2007; Dong et al., 2014). From this, the overall view emerging from the literature is that:
i Plants have evolved inducible defenses to deal with microbes that can break down physical barriers or take advantage of wounding events.
ii The proteins produced by the host during these defense responses are often secreted to combat infection.
iii That these weapons are not only directed against the pathogen cell itself (lytic enzymes) but also against pathogen proteins that are meant to facilitate infection (breaking down barriers or detoxify plant compounds).
These observations, in turn, support the view that there is an ongoing evolutionary conflict (driven by the life vs death stakes game) that drives the acquisition, adaptation, and selection of new molecular weaponry in both plants and pathogens. This is the reason that host-microbe interactions are often described as evolutionary arms races. The idea of co-evolutionary arms races also conforms with the “Red Queen Hypothesis” (Figure 3, a reference to the running red queen in “Alice through the Looking Glass” proposed by Leigh van Valen (1973)). Much like the red queen who must run on a moving chess board to stand still, organisms must continuously adapt (run) to survive (stand still) in their ever-changing natural environment (moving chess board).

Figure 3. The Red Queen explaining the running game to Alice. Image Source
Concluding thoughts
The notion of a co-evolutionary arms race only holds true if both the host and the pathogen are “running” to survive. Consistent with this idea, numerous studies have identified a range of evolutionary strategies, pathogens have adopted to keep up with evolving host plants. These fascinating and scintillating observations will be described in a future post (watch this space). Importantly though, the co-evolutionary arms race idea demands a re-think on how we cultivate large monoculture crops (elite varieties of a crop that are all genetically identical and are grown together in a field) and how this would impact these arms races. Again, this subject is worthy of its own post.
References:
· Boller T, Felix G. (2009) Annu Rev Plant Biol (2009) 60:379-406
· Choi, Hyong Woo & Kook Hwang, Byung. (2011). Systemic Acquired Resistance of Pepper to Microbial Pathogens. Journal of Phytopathology. 159. 393 - 400. 10.1111/j.1439-0434.2010.01781.x.
· Dodds PN, Rathjen JP. Plant immunity (2010). Nat Rev Genet 11(8):539-548
· Dong S, Stam R, Cano LM, Song J, Sklenar J, Yoshida K, Bozkurt TO, Oliva R, Liu Z, Tian M, Win J, Banfield MJ, Jones AM, van der Hoorn RA, Kamoun S. Science. (2014) Jan 31;343(6170): 552-5. doi: 10.1126/science.1246300.
· Freeman, B.C. and G.A. Beattie. 2008. The Plant Health Instructor. DOI: 10.1094/PHI-I-2008-0226-01
· Jones JDG, Dangl JL. Nature (2006) 444 (7117):323-329
· Schulze-Lefert P, Panstruga R. Trends in Plant Science 2011 16(3):117-125.
· Tian M, Win J, Song J, van der Hoorn R, van der Knaap E, Kamoun S. Plant Physiol. (2007) Jan;143(1):364-77. ·
· Van Valen, L. (1973). A new evolutionary law". Evolutionary Theory. 1: 1–30.
For further reading on this topic, I would recommend one of my other posts: @huitemae/pathogens-explored-features-that-set-microbial-parasites-apart-from-their-innocent-relatives
I hope you will find these pieces interesting. Your vote and follow help me open up the exciting world of plant biology and pathology to you.
Thank you @foundation for this fantastic SteemSTEM gif
